Дмитрий ГЛУХОВ, руководитель департамента КРС
Компания «Адиссео Евразия»
Протеин — это один из самых дорогих компонентов: затраты на него могут составлять от 35 до 55% от стоимости рациона. При этом эффективность использования белка в организме жвачных животных достаточно низкая (24–25%), она варьирует в широких пределах — от 10 до 40% (Calsamiglia et al., 2010). Этот показатель можно улучшить путем правильного подбора кормов и кормовых добавок и за счет грамотного балансирования рационов. В таком случае у специалистов по кормлению крупного рогатого скота возникают вопросы: какой протеин использовать и в каком количестве, как обеспечить максимальный уровень синтеза микробного белка в рубце и повысить эффективность использования транзитного (нерасщепляемого в рубце) протеина, чтобы сохранить здоровье животных, продлить их долголетие и получить качественную продукцию.
Протеиновое питание жвачных
Чтобы определить, какие источники протеина лучше использовать в кормлении коров, следует разобраться, как происходит усвоение кормового белка в организме жвачных, и понять, на какие показатели необходимо обращать внимание при составлении рационов.
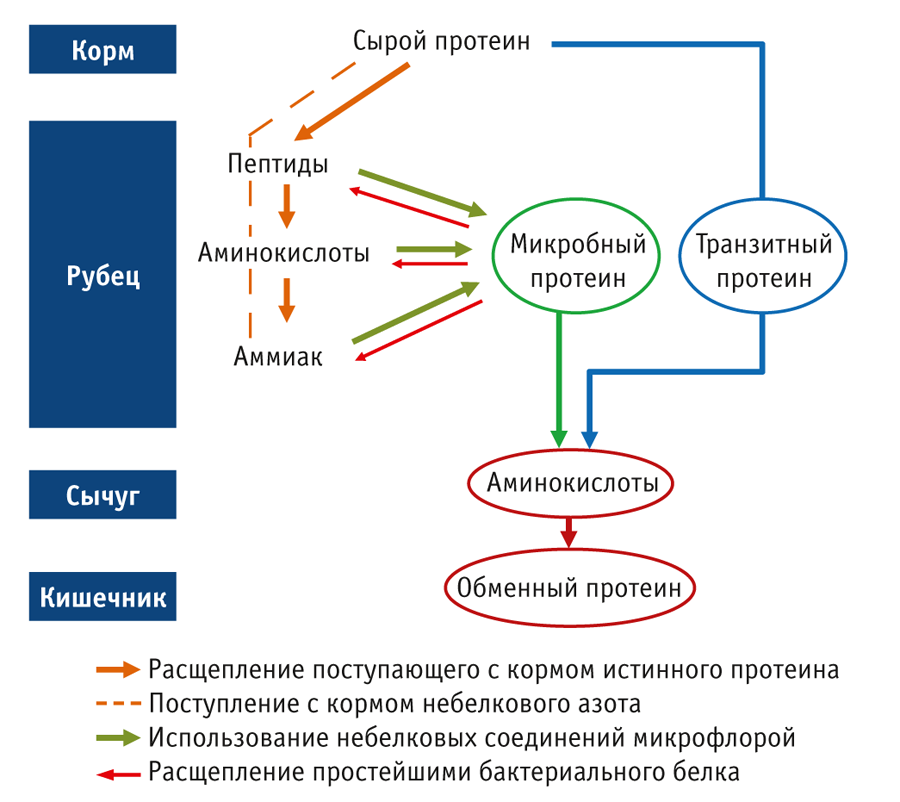
С кормом животное получает сырой протеин (СП). Это не только белок (или истинный протеин), но и небелковый азот, который представлен как более простыми органическими соединениями (например, пептиды, свободные аминокислоты, амиды, амины), так и неорганическими, в частности солями аммония. Часть истинного протеина доступна для рубцовой микрофлоры. Под действием протеолитических ферментов, вырабатываемых микроорганизмами, белки корма расщепляются до пептидов, аминокислот, а затем до аммиака (рис. 1). Все эти соединения наряду с поступившим с кормом небелковым азотом бактериальная микрофлора рубца использует для построения собственных клеток и роста.
До 50% бактерий служит питанием для простейших. Переваривая бактериальные клетки, они возвращают часть азота во внутреннюю среду рубца в виде все тех же небелковых соединений, которые затем снова становятся доступными для бактерий (Puniya et al., 2015). Этот процесс называют круговоротом или рециркуляцией азота в рубце.
Биомасса бактерий и простейших формирует 90–99% микробного протеина (Puniya et al., 2015) — основного и наиболее ценного источника белка для коровы. Микробный протеин, поступая в сычуг, расщепляется так же, как и транзитный протеин, до аминокислот, которые затем усваиваются в тонком кишечнике. В совокупности аминокислоты транзитного и микробного протеина составляют так называемый обменный протеин — белок, необходимый для поддержания жизнедеятельности и образования продукции (прирост живой массы и синтез молока). Этот показатель считается ключевым при составлении рационов. Он по‑разному именуется в различных системах расчета: MP (Metabolizable protein) в американских NRC и CNCPS, DVE (Darm Verteerbaar Eiwit) в нидерландской CVB, PDI (Protéine Digestible dans l’Intestin) во французской INRA или nXP (nutzbares Protein) в одноименной немецкой. При прогнозе молочной продуктивности необходимо учитывать показатель «содержание обменного протеина», а не «концентрация СП», который и заложен в программах, использующихся для расчета рационов.
Потребность коровы с удоями 25–30 кг молока в сутки в обменном протеине можно удовлетворить путем скармливания только качественных кормов и за счет максимальной выработки бактериями рубца микробного протеина. Если продуктивность животного достигает 30 кг и более, рубец зачастую неспособен обеспечить выработку необходимого количества обменного протеина. В результате этого возрастает роль транзитного протеина: чем выше продуктивность животного, тем больше транзитного протеина необходимо вводить в рацион. Однако для того, чтобы транзитный протеин полностью усваивался в организме коровы, его аминокислотный состав должен быть максимально близок к аминокислотному составу микробного белка. В силу этого при использовании транзитного протеина необходимо контролировать аминокислотный состав обменного протеина.
Оптимизация использования азота в рубце
Сколько СП нужно рубцовой микрофлоре? Максимального синтеза микробного белка достигают при концентрации 12–13% СП на 1 кг сухого вещества (СВ) рациона (с учетом того, что весь СП расщепляется в рубце). При среднем уровне СП 16–17% на 1 кг СВ на расщепляемый в рубце протеин должно приходиться 65–70%. Часто превышение этой нормы приводит к более серьезным последствиям, чем недостаток расщепляемого в рубце протеина.
Рубцовая микрофлора, в частности бактерии, использует для роста и размножения азот из небелковых соединений, которые либо поступают с кормом, либо образуются в рубце при расщеплении протеина. Основным источником азота служит аммиак — конечный продукт распада белка. Из него синтезируется от 50 до 80% микробного протеина. Однако бактерии могут использовать его в ограниченных количествах. Для максимально эффективного усвоения концентрация микробного протеина не должна быть слишком высокой — всего 5–6 мг/100 мл рубцовой жидкости (Satter, Slyter, 1974; Mehrez et al., 1977). Кроме того, аммиак легко всасывается через стенку рубца и при росте количества аммиака в рубцовой жидкости увеличивается и его концентрация в крови.
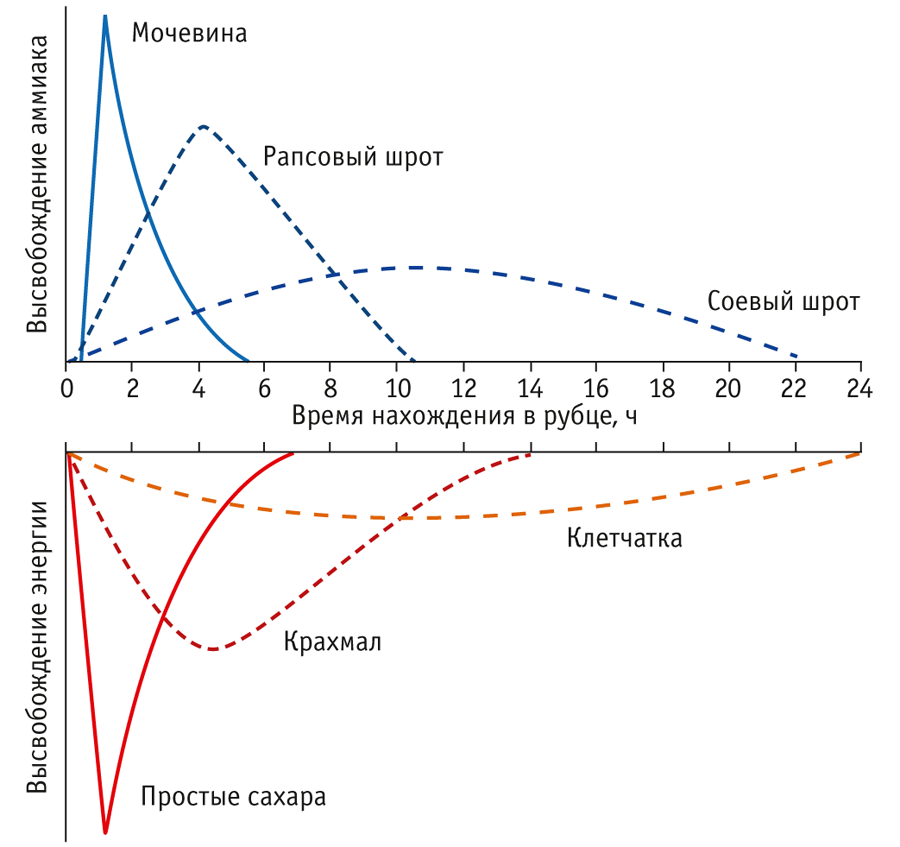
Пептиды и аминокислоты также относятся к важным источникам азота для микрофлоры: доля синтезируемого из них белка составляет 20–50%. Они служат источниками разветвленных углеродных цепей, необходимых для синтеза микробами аминокислот лейцина, изолейцина и валина, что особенно важно при использовании в рационах мочевины.
Помимо наличия источников азота для синтеза микробного белка необходимо наличие энергии, причем скорость высвобождения из кормов аммиака и энергии должна быть синхронизирована (рис. 2).
Последствия от неправильно сбалансированных по белку рационов
В случае поступления в рубец избыточного количества расщепляемого протеина или при недостатке энергии микрофлора не успевает использовать значительное количество высвобождаемого аммиака. Избыток аммиака негативно влияет как на рубец, так и на организм в целом. С ростом концентрации аммиака повышается и pH в рубце, что негативно сказывается на жизнедеятельности микрофлоры. Избыток аммиака также может ухудшить усвоение магния (Martens et al., 2018), а поскольку основная его часть усваивается именно в рубце, это может повлечь за собой ухудшение усвояемости кальция, что напрямую связано с усвоением магния. Аммиак через стенку рубца всасывается в кровь. Так как аммиак токсичен для организма и его накопление может вызвать серьезное отравление (Puniya et al., 2015), возникает необходимость в его обезвреживании. Происходит этот процесс в печени путем образования мочевины, причем затраты энергии в организме животного значительно возрастают: для образования 1 мг мочевины требуется 1,67 Дж энергии.
Из печени мочевина возвращается в кровяное русло, откуда легко проникает в прочие биологические жидкости. Часть мочевины со слюной поступает обратно в рубец (рециркуляция). При благоприятных условиях (когда в рубце нет избытка аммиака) мочевину используют бактерии для роста. В противном случае она будет расщеплена до аммиака (процесс происходит достаточно быстро и занимает от 30 минут до 2 часов), снова всосется в кровь и процесс рециркуляции возобновится.
Скармливание избыточного количества расщепляемого в рубце протеина может привести как минимум к трем негативным последствиям: нарушению работы рубца, снижению усвояемости магния и, опосредованно, кальция, а также к увеличению затрат энергии. Уже это отрицательно сказывается на здоровье и продуктивности коров, особенно в начале лактации. Однако и образующаяся в итоге мочевина может также негативно повлиять на состояние животных и качество молока. В частности, при повышении уровня мочевины у коров ухудшается функция воспроизводства.
Поскольку мочевина легко проникает во все органы и ткани, она способна влиять на рН внутренней среды матки, а также на концентрацию магния, калия, фосфора и цинка в маточной жидкости. В результате снижаются и оплодотворяемость (за счет уменьшения жизнеспособности попадающих в матку сперматозоидов), и выживаемость эмбрионов на ранних стадиях развития, так как в первые 26–28 дней стельности зародыш не прикреплен к стенке матки (табл. 1). Вместе с тем мочевина негативно влияет на уровень прогестерона, то есть напрямую сказывается на стабильности беременности. В результате уменьшается процент плодотворно осемененных животных при первом и втором осеменении, что влечет за собой экономические потери (дополнительные расходы, связанные с покупкой спермодоз, увеличением продолжительности сервис-периода и интервалов между отелами).

Помимо этого с ростом уровня мочевины в крови растет ее концентрация в молоке. Это в свою очередь приводит к тому, что концентрация истинного протеина (казеин и сывороточный белок) в молоке снижается и оно становится менее пригодным для приготовления сыра или творога, то есть выход конечного продукта при переработке существенно уменьшается.
В то же время мочевина — хороший индикатор правильности протеинового кормления. В частности, можно использовать такие показатели, как АМП и АМК. Они тесно связаны друг с другом и рассчитываются по формуле
АМП, мг/100 мл = (1,021 × АМК) + 0,399.
Еще один хороший индикатор эффективности белкового обмена в организме коров — АММ (Roseler et al., 1993). Этот показатель может использоваться для оценки правильности кормления в силу того, что он напрямую коррелирует с АМК, так как мочевина легко переходит из крови в другие ткани (Melendez et al., 2000). Взаимосвязь между показателями рассчитывают по формуле
АММ, мг/100 мл = (0,620 × АМК) + 4,75.
Целевое значение концентрации АММ составляет 8–10 мг/100 мл (Schwab, 2015), что соответствует 171–214 мг/л мочевины.
В каких же случаях может возникать избыток расщепляемого в рубце протеина? Чаще всего это происходит из‑за скармливания животным кормосмесей, содержащих более 17% СП в 1 кг СВ. Балансируя рационы, специалисты повышают содержание протеина, стараясь увеличить в кормосмеси концентрацию обменного протеина до уровня, необходимого для получения плановых показателей по надоям. При этом не учитывают общее содержание протеина в корме и забывают о таком показателе, как азотный баланс рубца. Его названия в разных системах различаются (например, Rumen NH3 balance, OEB, RNB), так же как и способы расчета, но в любом случае именно такой показатель служит ключом для правильного расчета как количества азота в рубце, так и его соотношения с энергией.
Игнорирование этого показателя — самая распространенная ошибка. Иногда она возникает из‑за невнимательности специалиста по кормлению, а иногда из‑за стремления сэкономить. Так, в некоторых хозяйствах часть соевого или рапсового шрота, содержащих в среднем 30% нерасщепляемого в рубце протеина (NRC, 2001), заменяют на подсолнечный, бОльшая часть белка (не менее 84%) в котором расщепляется в рубце, либо на мочевину, которая, как известно, под действием ферментов микрофлоры очень быстро распадается до аммиака. Вследствие такой замены нарушается азотный баланс рубца.
Помимо высокого уровня СП причиной нарушения азотного баланса может стать недостаток энергии в рубце или отсутствие синхронизации высвобождения азота и энергии (об этом написано выше). Такая ситуация может возникнуть в случае, когда «быстрый» азот, например из той же мочевины, не сбалансирован необходимым количеством «быстрой» энергии, то есть сахаров. В результате даже при оптимальном уровне СП в рационе существенная часть образующегося в рубце аммиака не будет использоваться микрофлорой.
Избыток расщепляемого в рубце протеина также часто встречается у животных в тех хозяйствах, где в качестве основного корма используют сенаж и сено из люцерны или других бобовых, а в качестве дополнительного источника протеина — жмых или шрот из подсолнечника или рапса.
Как исправить ситуацию?
Если в рационе концентрация СП превышает 17% в 1 кг СВ, следует в первую очередь уменьшить его количество, причем сделать это желательно за счет использования кормов с наиболее высоким содержанием расщепляемого в рубце протеина. Далее необходимо попытаться нормализовать азотный баланс рубца, то есть обеспечить как правильное количественное соотношение источников азота и энергии, так и синхронное высвобождение азота и энергии.
Если азотный баланс по‑прежнему высокий, нужно заменить часть имеющихся источников белка кормами с более высоким содержанием транзитного протеина, такими как соевый шрот, сухая послеспиртовая барда, пивная дробина, кукурузный глютен или так называемый защищенный протеин. Можно снизить расщепляемость протеина и рециркуляцию азота внутри рубца путем ввода в рацион танинов и сапонинов.
И те и другие присутствуют во многих растениях и способны влиять на расщепляемость протеина и активность микрофлоры.
При добавлении в кормосмесь сапонинов снижается активность простейших (Patra, Saxena, 2009), в результате чего уменьшается поглощение ими бактерий и количество циркулирующего в рубце азота (см. рис. 1). Благодаря этому доля бактериального белка в микробном протеине возрастает, что имеет определенные преимущества: бактерии размножаются быстрее простейших и при этом живут гораздо меньше, то есть таким образом обеспечивается более оперативное поступление преобразованного азота корма в сычуг. Иными словами, животное получает больше микробного белка.
Ввод в рацион танинов позволяет преобразовать часть расщепляемого в рубце белка в транзитный протеин (Frutos et al., 2004; Naumann et al., 2017; Nawab et al., 2020). Происходит это благодаря тому, что танины способны связываться с протеином корма, делая его недоступным для ферментов микрофлоры. Попадая в кислую среду сычуга, эти соединения разрушаются, и белок становится доступным для переваривания и последующего усвоения.
Помимо этого танины способны частично уменьшать ферментативную активность некоторых бактерий, за счет чего снижается расщепление пептидов и аминокислот в рубце до аммиака. Таким образом, при воздействии танинов не только снижается выработка аммиака, но и увеличивается количество аминокислот, получаемых животным из белка кормов.
Танины и сапонины можно использовать как по отдельности, так и в комбинации. Эффективность применения смеси сапонинов и танинов в кормлении крупного рогатого скота подтверждена на практике. Данные экспериментов показали, что при включении такой смеси в рацион (50 г на голову в сутки) снизился уровень мочевины в крови животных с 32 до 27 мг%, среднесуточная продуктивность выросла в среднем на 1 кг на голову в сутки (с 32,8 до 33,8 кг), а кроме того, улучшилось качество молока (массовая доля белка увеличилась с 3,32 до 3,37%, жира — с 3,74 до 3,84%).
Однако стоит помнить, что сапонины и танины — не волшебная таблетка. Их целесообразно применять только в тех случаях, когда по тем или иным причинам нет возможности снизить количество расщепляемого в рубце белка или оптимизировать азотный баланс.
Повышение эффективности транзитного протеина
Напомню, что при продуктивности менее 30 кг молока в сутки коровы способны практически полностью удовлетворять свою потребность в обменном протеине за счет белка, синтезируемого микрофлорой рубца. Микробный белок — наиболее полноценный для животных с точки зрения аминокислотного состава — организм использует максимально эффективно.
При продуктивности свыше 30 кг, как правило, возникает необходимость в применении источников транзитного протеина. Задача специалистов по кормлению — обеспечить животных транзитным протеином, аминокислотный состав которого будет максимально близок к аминокислотному составу микробного белка. В этом случае не возникнет большого количества неиспользуемых аминокислот, которые организму животного придется утилизировать.
Как известно, утилизация «лишних» аминокислот происходит большей частью в печени, а конечным продуктом утилизации будет аммиак, который необходимо обезвредить, превратив в мочевину. Таким образом, скармливание большого количества несбалансированного по аминокислотному составу транзитного протеина может привести к тем же последствиям, что и излишек расщепляемого в рубце белка (увеличение нагрузки на печень, дополнительные энергозатраты, ухудшение здоровья и воспроизводительной способности).
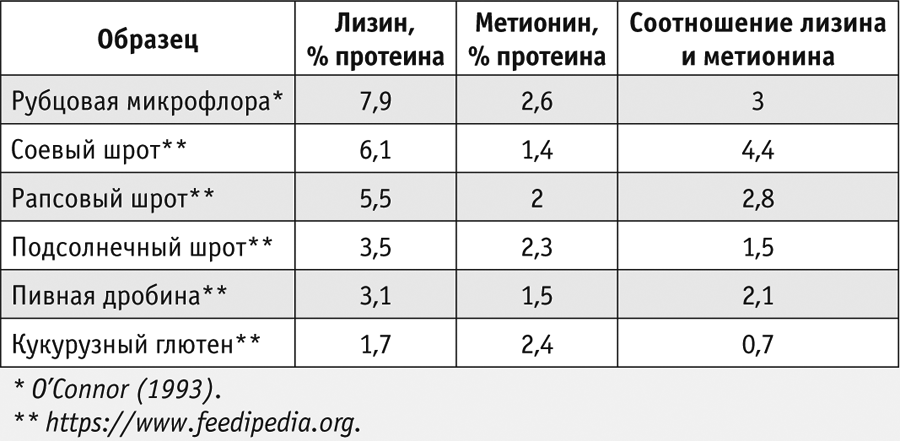
При использовании источников транзитного протеина растительного происхождения, как правило, возникает дефицит двух аминокислот: метионина и лизина (табл. 2), то есть именно эти две аминокислоты будут лимитировать использование обменного белка и, соответственно, образование продукции.
В этом случае возникает необходимость в использовании отдельных аминокислот в защищенной от распада в рубце форме, причем нормировать их нужно так же, как и другие питательные вещества. Для защищенных метионина и лизина нет фиксированных норм ввода, для каждого рациона их количество рассчитывают индивидуально в зависимости от применяемых источников протеина. Например, в рационы с высоким содержанием соевого шрота обычно требуется дополнительно вводить только метионин. При полной или частичной замене сои рапсом необходимо добавлять еще и лизин. В кормосмеси с бардой или дробиной дефицит лизина будет более выраженным.
Итак, как правильно сбалансировать рацион, чтобы повысить эффективность использования протеина в организме жвачных? Нужно оптимизировать рубцовое пищеварение путем грамотного балансирования рационов, во‑первых, по количеству расщепляемого протеина, во‑вторых, по соотношению азота и энергии (азотный баланс рубца). Необходимо также синхронизировать высвобождение азота и энергии из их источников. Это позволит получить максимальное количество микробного протеина — самого дешевого и при этом наиболее полноценного источника обменного протеина для коров. При невозможности нормализовать азотный баланс можно попытаться оптимизировать рубцовое пищеварение путем включения в рацион танинов и сапонинов.
Специалисты рекомендуют при вводе в кормосмесь источников транзитного протеина, особенно растительного происхождения, использовать защищенные метионин и лизин. Дозировку каждой из аминокислот необходимо рассчитывать исходя исключительно из структуры рациона и качества кормов, применяемых на предприятии.
Вопросы автору можно направлять на e-mail: Dmitry.Glukhov@adisseo.com